Biologie Cellullaire :
Chapitre 5 : Le Rein/Appareil Urinaire.

Télécharger le document sur le serveur FTP ou sur le serveur web (HTTP).
I Généralités.
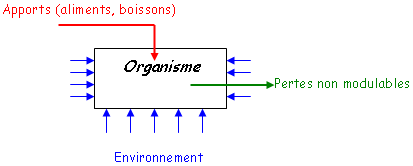
A Régulation.
Un organisme, par ses apports alimentaires, son environnement, par les pertes non modulables (eau pendant la transpiration) va devoir développer un système de régulation de son milieu intérieur : l’homéostasie.
1 Homéostasie hydro-électrolytique.
Le rein joue un rôle dans la régulation du capital hydrique, minéral et dans le maintien de la stabilité du pH dans le sang et le milieu intérieur.
2 Evacuation des déchets cataboliques.
Parmi les déchets du catabolisme, on trouve les déchets azotés comme NH3 (ammoniac) qui est toxique. Il va y avoir élimination de NH4+ (ammonium) ou d’urée ou d’acide urique. Cette régulation reflète la fonction exocrine du rein.
3 Régulation de la pression artérielle.
Le rein joue aussi un rôle endocrine, comme avec l’EPO ou le système rénine/angiotensine pour la régulation de la pression artérielle.
B Le rein de vertébrés.
Le rein est un organe massif, organisé de très nombreuses unités fonctionnelles : le néphrons. Le néphron est constitué de deux parties.
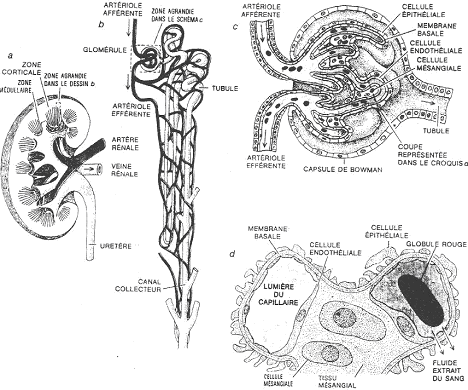
1 Le néphron.
a Le corpuscule.
Le corpuscule est l’endroit où se forme l’urine primitive, par filtration du sang. Le corpuscule est le glomérule accolé à la capsule. Le glomérule est une pelote de capillaires.
Le sang arrive par l’artère rénale et va être ultra-filtré. Le sang ressort par une veine efférente.
La vasa recta est un système circulatoire à contre courant.
b Les tubes.
Les tubes ont un trajet contourné. Ils ont pour fonction de conduire l’urine de la capsule jusqu’à l’extérieur (parfois, il n’y a pas de vessie).
Les tubes vont modifier l’urine en fonction du type cellulaire présent.
c L’organisation du néphron.
L’organisation du néphron est presque toujours la même. Les groupes les moins évolués ont les néphrons les moins évolués.
Le néphron ouvert est un glomérule pendu dans la cavité cœlomique, à son contact direct. Le sang ultra-filtré et l’urine transitent par la cavité cœlomique puis sont récupérés par un canal, le néphrostome, à épithélium simple, cuboïdal, cilié. On trouve cette organisation chez les alevins de poissons ou chez les larves de certains amphibiens.
Au cours de l’évolution, les néphrons se sont fermés. L’urine passe directement dans des tubes : ce sont les néphrons fermés. C’est le cas le plus général, sauf chez certains poissons marins qui ont des néphrons aglomérulés.
Dans ce cas, on n’a uniquement des tubes où se font les échanges, modifiant l’urine éliminée par un canal collecteur. è C’est une adaptation à des conditions de vie en forte osmolarité (à problème de la rétention d’eau).
Le néphron est toujours organisé en deux parties mais la subdivision des tubes peut être différente :
- Chez les non mammifères et non oiseaux, on a un corpuscule, un tube contourné proximal, un tube contourné distal et un canal de Wolf (évacuateur).
- Chez les mammifères et oiseaux, on a le corpuscule, le tube contourné proximal, le segment intermédiaire (anse de Henlé chez les mammifères), le tube contourné distal et des canaux collecteurs qui font déboucher l’urine dans un uretère secondaire. Il y a mise en place d’un canal spécifique à l’évacuation de l’urine.
L’apparition de ce segment intermédiaire régionalise le rein : on distingue maintenant une région corticale (le cortex) qui contient les corpuscules, et, une région médullaire (la médulla) qui regroupent les tubes contournés distaux et les anses de Henlé.
2 Le développement embryonnaire.
Le rein se met en place à partir d’une ébauche métamérisée. Cette disposition segmentaire implique un faible nombre de segments. La différenciation est progressive, d’avant en arrière et par étapes.
Il va y avoir mise en place d’un pronéphros (le rein définitif de poisson et des amphibiens). La disposition segmentaire va disparaître. Le nombre de néphrons augmente et l’urine est évacuée par le canal de Wolf (urine et produits sexuels).
Cette structure est temporaire chez les amniotes.
Il y a apparition du métanéphros qui voit se former un uretère secondaire spécialisé.
II Le rein de mammifères.
Les néphrons sont fermés, glomérulés et régionalisés. La circulation des vaisseaux sanguins souligne la ligne de séparation entre le cortex et la médullaire.
Il existe deux types de néphrons : les néphrons corticaux et les néphrons juxta-glomérulaires. Le fonctionnement de ces deux types est différent. Une différence morphologique importante est la différence de la taille de la anse de Henlé.
A Le corpuscule.
Le glomérule est contenu dans la dépression d’une capsule. En coupe, on voit dans le corpuscule, de nombreux capillaires sanguins : le glomérule. La capsule est constituée de deux feuillets : le feuillet pariétal (est bordé par un épithélium simple, squameux) et le feuillet viscéral (à épithélium formé de podocytes).
Les podocytes sont caractérisés par leurs prolongements cytoplasmiques se ramifiant, appelés, les pédicelles. Ces derniers s’appliquent étroitement autour des capillaires des glomérules.
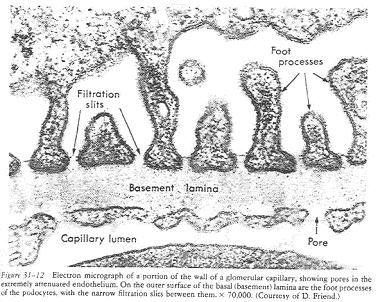
Entre les pédicelles et l’endothélium (des capillaires), on a les deux lames basales qui ont fusionné. Le sang arrive dans les capillaires sanguins, traverse cette barrière (de filtration) constituée d’un endothélium (fenêtré), d’une lame basale et d’un pédicelle.
L’espace ménagé entre les pédicelles s’appelle les « fentes de filtration ». La filtration du sang est un mécanisme passif se faisant grâce à la pression sanguine existant dans les capillaires. La pression dans les glomérule est forte (» pression aortique). Une diminution de cette pression entraîne une diminution de la filtration et une diminution de la quantité de l’urine.
La pression provoque le passage du sang vers l’espace de Bowman. Seul le plasma passe dans l’espace de Bowman (sans les grosses protéines).
Le tamis moléculaire vrai est la lame basale. Certains diabètes rénaux montrent une anomalie de cette lame basale. Au niveau des pores de l’endothélium, on observe un diaphragme (glycocalix anionique). Ils ne servent qu’à ralentir le passage des molécules.
Un autre type cellulaire est présent : ce sont les cellules mésengiales. Ce sont l’équivalent des macrophages et elles ont pour rôle principal, la dégradation des constituants de la lame basale. Elles permettent ainsi le renouvellement des constituants de cette lame basale. Le renouvellement est assuré par les podocytes.
Les cellules mésengiales servent aussi à l’élimination des grosses protéines (Ig) coincées dans la lame basale. On pense que ces cellules ont des caractéristiques communes aux péricytes (propriété contractile pour réguler le flux sanguin).
B Les tubes urinaires.
Les tubes urinaires sont les tubes contournés proximaux et les tubes contournés distaux. Ils conduisent l’urine mais vont aussi la modifier. Trois mécanismes entrent en jeu pour la modification : la diffusion, la sécrétion tubulaire, la réabsorption spécifique.
Les sécrétions sont les produits synthétisés par les cellules bordantes. Chaque tube a des caractéristiques reflétant ses cellules bordantes.
1 Le tube contourné proximal.
Le tube contourné proximal est bordé par un épithélium simple (palissadique ou cuboïdal), avec une bordure en brosse. Ces microvillosités montrent une augmentation de la surface d’échange. Ce tube peut aussi faire de l’endocytose (99% des acides aminés sont réabsorbés par ce tronçon ainsi que 100% des sucres).
Les cellules sont caractérisées par les microvillosités, des vésicules d’endocytose et au pôle basal, de nombreux replis membranaires (avec des mitochondries en alignement).
2 La anse de Henlé ou segment intermédiaire.
On trouve ces parties dans la médullaire.
La taille de ces anses varie en fonction de l’habitat de l’animal. Elle est courte quand l’animal vit près de l’eau et plus longue s’il vit dans des régions plus arides. Elle permet une lutte contre la déshydratation.
La anse de Henlé joue un rôle dans la concentration de l’urine.
La branche descendante est perméable à l’eau et aux sels. La branche ascendante est imperméable à l’eau et peut faire sortir activement les sels.
Si des sels sont dans le milieu interstitiel, ils entrent dans la branche descendante, donc, de l’eau sort. En descendant, l’urine va se concentrer. Elle se diluera en remontant, sous les mouvements d’eau : il se met en place un gradient cortico-médullaire. La concentration du tissu interstitiel s’accroît en allant vers le bas de la anse.
A la suite de cette anse, on trouve le tube contourné distal et le tube collecteur ; ils sont perméables à l’eau à certaines conditions. Comme le milieu interstitiel est concentré, le tube collecteur perd de l’eau et donc, concentre l’urine.
La anse de Henlé ne fait qu’augmenter la pression osmotique du tissu interstitiel pour que le tube collecteur perde beaucoup d’eau è économie d’eau. Il faut de l’ADH pour que le système fonctionne. Comme le tube collecteur est perméable à l’urée, il y a augmentation de la pression osmotique du tissu interstitiel (accroissement du phénomène).
La branche descendante diminue de diamètre (devient grêle) : il y a une surpression hydrostatique qui provoque la sortie d’eau.
La vasa recta est un système qui récupère l’eau qui sort des tubes : elle maintient le gradient.
La anse de Henlé, chez les mammifères, est bordée par un épithélium simple, squameux.
3 Le tube contourné distal.
Le tube contourné distal est bordé d’un épithélium simple, sans microvillosité, ni entérocyte, ni aldostérone. Il présente quelques replis membranaires avec alignements de mitochondries.
Le tube est capable de former la macula densa.
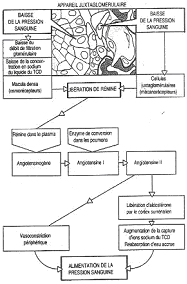
Les cellules du tube contourné distal sont capables, pour certaines, de se différencier en macula densa. Ces cellules deviennent des chémorécepteurs (sensibles à la concentration de sodium dans le tube contourné distal). La macula densa est au contact des artérioles afférentes et efférentes.
Certaines cellules musculaires lisses, autour de l’artériole afférente, se différencient en cellules à rénine. Ces cellules sont des barorécepteurs.
C’est l’appareil juxta-glomérulaire.
Quand la pression sanguine diminue dans l’artériole, ces cellules libèrent de la pro-rénine dans le sang. Elle va être clivée et donnera de la rénine.
Pour l’interaction avec l’angiotensine, voir cours de BGU03 (endocrinologie).
Une diminution de la concentration en sodium dans le tube contourné distal stimule la macula densa et stimule les cellules à rénine.
4 Le tube collecteur, l’uretère et la vessie.
Le tube collecteur a un épithélium simple, palissadique, sans bordure en brosse, sensible à l’ADH (réabsorption d’eau). Il permet un ajustement final de l’urine (concentration, volume).
L’urine passe dans l’uretère bordé d’un épithélium de transition (stratifié). Le même type d’épithélium est présent dans la vessie. Les cellules, au pôle apical, on une région de membrane plasmique semblant rigide : la membrane plasmique apicale possède des phospholipides avec beaucoup de cholestérol, ce qui donne cette rigidité membranaire. à Les régions rectilignes sont séparées par des régions plus souples.
Dans le cytoplasme, on a une mise en réserve de membrane plasmique (sous forme d’aiguilles à cause de la rigidité) qui pourra être réutilisée au pôle apical. C’est une adaptation des surfaces membranaires.
La vessie vide montre un épithélium épais alors que pleine, il semble fin. C’est une adaptation pour l’élasticité de la vessie.
III Rein et appareil excréteur d’oiseau.
Les néphrons sont fermés, glomérulés et régionalisés.
Le tube contourné proximal est comme celui des mammifères, ainsi que le tube contourné distal. Le segment intermédiaire est différent : l’épithélium est simple, cuboïdal. Les tubes collecteurs ont un épithélium palissadique.
Chez les oiseaux ont a des paquets de médullaire mélangés à du cortex.
On trouve un uretère (secondaire) mais pas toujours de vessie. Cet uretère est bordé d’un épithélium pseudostratifié, très haut, riche en cellules à mucus, avec souvent, du tissu lymphoïde.
Il y a une lutte contre la déshydratation par les phanères avec les plumes. Les oiseaux ont une urine riche en acide urique qui peut être évacué dans un faible volume d’eau : l’urine est quasiment solide.
IV Rein de batraciens.
Le rein de batraciens a des néphrons fermés, glomérulés et alignés (pas de segment intermédiaire). On voit souvent deux lignes de néphrons : on peut reconnaître les tubes contournés proximaux et tubes collecteurs.
Les tubes contournés proximaux et les tubes contournés distaux sont similaires à ceux des mammifères.
Le pôle urinaire est cilié.
V Rein de reptiles.
Le rein de reptile est une association de rein et de tissus génital. Les néphrons sont classiques ainsi que les tubes contournés proximaux, les tubes contournés distaux et les tubes collecteurs.
VI Rein de poisson.
Le rein de poisson occupe presque toute la longueur de l’animal (dans la cavité abdominale).
On trouve un rein céphalique (sans fonction urinaire) qui est un organe lymphoïde, et un rein postérieur, à fonction urinaire.
Les néphrons sont fermés, glomérulés et dispersés. Les tubes contournés proximaux sont classiques. Les tubes contournés distaux et les tubes collecteurs ont un épithélium simple cuboïdal.
Il n’y a pas de régionalisation du rein, ni d’alignement. On ne peut pas faire la distinction entre les tubes contournés distaux et les tubes collecteurs.
Le canal de Wolf ou uretère primitif a un épithélium pseudostratifié avec des cellules à mucus. Les corpuscules et les tubes baignent dans du tissu hématopoïétique. On voit aussi des cellules équivalent à des macrophages (tissu atrocytaire).
On trouve souvent une glande régulant la calcémie (corpuscule de Stanius) ainsi que des follicules thyroïdiens fonctionnels.
Le rein de poisson est un organe composite, comprenant différents types cellulaires et assurant diverses fonctions. Il ne présente pas de régionalisation.
Aucun commentaire jusqu'à présent.