Biologie du Développement :
Introduction à la Biologie du Développement.
Télécharger le document sur le serveur FTP ou sur le serveur web (HTTP).
Introduction.
L’embryologie constitue l’étude du développement embryonnaire de l’œuf unicellulaire à un individu pluricellulaire autonome. On passe selon des séquences ordonnées que l’on peut assimiler à un programme du développement de l’état relativement simple d’un œuf, aux structures relativement complexes d’un adulte.
L’activation de l’ovocyte 2 qui entraîne la formation de l’œuf peut être due à une fécondation ou à un développement parthénogénique (sans fécondation).
Le développement embryonnaire comprend de grandes étapes.
I L’œuf donnera le zygote.
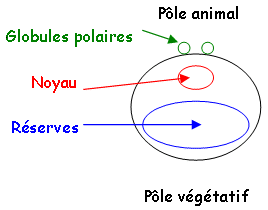
L’œuf est totipotent et relativement simple. Il résulte de la fécondation de l’ovule, par le spermatozoïde. Au cours des deux divisions méiotiques, il y a migration des deux globules polaires vers le pôle supérieur de l’ovocyte (le pôle animal). Le noyau subit des divisions méiotiques et sera excentré dans la cellule.
<>Le pôle animal est composé d’un noyau et de globules polaires. Le pôle végétatif constitue des réserves. L’axe entre le pôle animal et le pôle végétatif est appelé l’axe antéropostérieur. Le pôle animal donnera la tête alors que le pôle végétatif donnera la queue. Le devenir de ces deux pôles est déterminé durant l’orogenèse.
Les zygotes ont plus ou moins de réserves. Selon les réserves en vitellus, on a :
- les zygotes alécithes : ils sont sans réserve et se trouvent chez les mammifères ;
- les zygotes oligocithes : ils ont peu de réserves et sont trouvés chez les oursins ;
- les zygotes hétérolécithes : ils ont beaucoup de réserves, on les trouve chez les amphibiens comme les anoures ;
- les zygotes télolécithes : c’est le jaune de l’œuf de poule. Le blanc donne l’albumen.
Après la fécondation : cet œuf va se segmenter par le phénomène de » segmentation « . Cette action va dépendre de la quantité des réserves.
II La segmentation.
Les cellules vont se diviser tout en restant jointives : ce sont les cellules filles. Elles donneront les blastomères (la partie du germe). Les cycles cellulaires vont se dérouler rapidement et sont synchrones au début puis deviendront asynchrones par la suite : on obtient alors une morula (en forme d’une mûre).
La segmentation peut-être totale (selon les réserves) :
- égale : les cellules sont de même taille (pendant les trois premiers cycles cellulaires chez l’oursin, et pendant les deux premiers cycles cellulaires chez les amphibiens).
- inégale : les blastomères sont de tailles différentes : on a une orientation des cellules et une apparition de fuseaux (excentrés). À cause de l’orientation en fuseaux, on aura une petite cellule ou des grosses (des micros ou des macromères).
Remarque : l’œuf garde durant toute cette phase le même diamètre et la même taille. Seul le nombre de cellules augmente.
La segmentation partielle : Elle a lieu chez les oiseaux (les télolécithes). Elle se limite à une petite région de l’œuf (territoire cytoplasmique), c’est une segmentation discoïdale, elle se limite à un disque : c’est la cicatricule du pôle animal.
Le nombre de cellules va augmenter :
- de 8 à 16 cellules, on a une morula.
- de 16 à 32 cellules, on a un blastocèle (cavité ) qui apparaît. C’est une blastula.
Le cytoplasme de l’œuf fécondé est hétérogène car la fécondation a pour effet de provoquer des remaniements cytoplasmiques avec des localisations préférentielles de molécules (ARN ou protéines). On a des déterminants moléculaires qui préféreront la zone ventrale ou dorsale dans le futur de l’œuf : on parle de détermination.
Au cours des divisions successives, les potentialités vont se réduire. Les blastomères vont subir une étape de détermination et deviendront pluripotents puis ces cellules se différencient et deviennent unipotentes.
Les mécanismes mal expliqués dans la détermination et la différenciation sont très complexes et font intervenir des facteurs de régulation des gènes et des facteurs intervenants dans la communication cellulaire.
A les facteurs de régulation des gènes :
Les étapes de détermination cellulaire peuvent apparaître très tôt dans le développement, dès la segmentation (phase de clivage de l’œuf). Les facteurs internes sont apportés par la mère au cours de l’orogenèse (ARN, protéines, enzymes, facteurs de croissance) et stockés dans l’ovocyte. La segmentation répartis ces facteurs de façon inégale dans les blastomères qui héritent de potentialités distinctes dont le rôle du génome maternel sur le développement de l’œuf dans le cas de certaines mutations apportées par la mère : ce sont des mutations à effet maternel. Ces changements de génome vont affecter certaines molécules indispensables au développement de l’embryon.
La détermination cellulaire n’est visible ni morphologiquement ni fonctionnellement. Elle prépare la différenciation cellulaire qui entraîne des manifestations morphologiques et fonctionnelles.
Il faut attendre plusieurs cycles (une dizaine) pour que s’exprime le génome du zygote. C’est la transition blastuléenne : on passe d’une expression maternelle des gènes à une expression zygotique. Les noyaux vont transcrire leur propre ADN. On peut donc dire qu’il y a deux programmes : un programme maternel et un programme zygotique.
B Les facteurs intervenants dans la communication cellulaire:
Ces facteurs activent des gènes qui vont donner des molécules transloquées aux membranes ou déposées sur une matrice extracellulaire permettant aux cellules de se reconnaître. On a ainsi l’établissement d’un territoire cellulaire ayant des potentialités différentes, appelé territoire présomptif de trois types différents durant la segmentation :
- ectoblaste.
- mésoblaste.
- endoblaste.
Chaque territoire est déterminé pour donner un certain type de tissus.
La segmentation aboutit à la répartition différentielle du contenu de l’œuf et à la constitution d’ensembles cellulaires à potentialités distinctes.
III La gastrulation.
Il y a mise en place de la blastula (le futur estomac).
Les cellules vont migrer par une phase de migration cellulaire. On a des mouvements morphogénétiques : la morphogenèse. C’est elle qui va modeler l’embryon.
La gastrulation est une phase dynamique.
Cette morphogenèse est caractérisée par des déplacements cellulaires qui vont concrétiser la mise en place des territoires présomptifs. C’est le résultat de mouvements cellulaires de certaines régions de la blastula, de l’organisation de populations cellulaires définies qui constitueront l’ectoderme, le mésoderme et l’endoderme, du positionnement de ces feuillets les uns par rapport aux autres.
Tous ces mécanismes conduisent à l’établissement de nouvelles interactions cellulaires qui préparent l’embryon à la phase d’organogenèse. Certaines de ces interactions sont des inductions.
L’ectoderme donne la plaque neurale grâce à ces interactions. Il faut pour cela que l’ectoderme soit compétent à recevoir des informations pour activer certains gènes. Les tissus inducteurs et compétents sont limités dans le temps. À certain stade, l’ectoderme peut être ou non compétent. Un tissu compétent peut devenir inducteur à certains stades.
Remarque : on a une cascade d’inductions. Cette induction peut être une levée d’inhibition.
Ces mécanismes font appel soit à des molécules sécrétées soit à des récepteurs membranaires qui transmettent des signaux jusqu’au gènes et permettent l’activation d’intégrines avec une cascade de phosphorylation et, dès qu’il y a contact, il y aura activation.
IV L’organogenèse.
La détermination cellulaire des territoires présomptifs va engendrer la formation des organes et leur différenciation cellulaire.
À l’organogenèse est liée la morphogenèse : le modelage du corps de l’embryon. De cette phase va sortir un organisme doté d’une certaine autonomie. Il aura soit le même plan d’organisation que l’adulte, soit une organisation différente avec un stade larvaire puis, une métamorphose et enfin, dans différent phylum des structures embryonnaires transitoires qui sont constituées puis qui vont disparaître et vont permettre le développement de l’embryon.
Parmi les annexes embryonnaires, on trouve :
- l’amnios (cavité amniotique).
- allantoïde.
- des vésicules vitellines qui donneront la première lignée sanguine.
trés interessant.
Bonjour, merci pour ce super cours je n’ai juste pas compris ce passage dans I/ L’œuf donnera le zygote.
Le pôle animal donnera la tête alors que le pôle végétatif donnera la queue. Le devenir de ces deux pôles est déterminé durant l’orogenèse.
Quel est le rapport avec l’orogénèse ? (est ce bien l’ensemble des mécanismes de formations des montagnes ?)
Coridalement