Physiologie Végétale :
Partie 1 : nutrition et métabolisme.
Chapitre 4 : La nutrition en eau.

Télécharger le document (toute la partie 1) sur le serveur FTP ou sur le serveur web (HTTP).
I L’eau du sol.
A Liaisons de l’eau.
L’humidité est exprimée en pourcentage de la masse. C’est une notion vague, car un sable, à 10% d’eau paraît humide alors qu’une tourbe à 50% d’eau sera sèche.
L’eau peut-être libre ou liée aux constituants du sol. On trouve l’existence de plusieurs forces : osmotique, capillaire (tension superficielle), électrostatique (imbibition).
B Potentiel hydrique et succion.
ψ : c’est la force d’attraction entre l’eau et le sol. Le potentiel hydrique est égal mais de signe opposé à l’énergie qu’il faut appliquer pour libérer 1 gramme d’eau. La valeur de ce potentiel est toujours inférieure à 0. Plus cette valeur est basse, plus les liaisons eau/sol sont fortes. Quand un sol se dessèche, ψ va baisser. La succion représente la force d’attraction exercée du sol sur l’eau (cette succion est considérée comme une pression, ainsi que le potentiel hydrique).
C La capacité de rétention.
C’est la quantité d’eau (en gramme), contenue dans 100g de sol après centrifugation à 500G ou après drainage.
D Le point de flétrissement.
Le point de flétrissement permanent est le taux d’humidité d’un sol, pour lequel, une plante flétrie irréversiblement (sable 5%, limon 12%, argile 27%, tourbe 50%).
E Réserve utilisable.
La réserve utilisable est la mesure de la quantité d’eau utilisable par la plante. C’est la différence entre la capacité de rétention et le point de flétrissement initial. Cette réserve représente généralement la moitié de la capacité de rétention. Un sol léger aura besoin d’un arrosage fréquent alors qu’un sol lourd n’en aura pas besoin.
II L’eau dans la plante.
A La teneur en eau des végétaux.
Leur grande vacuole leur sert de réservoir d’eau.
Dans la plante, le xylème et le phloëme sont les vaisseaux qui conduisent les deux sèves. Le xylème est un ensemble de tissus morts, où circule la sève brute (eau +sels minéraux). Le phloëme est composé de tissus vivants où circule la sève élaborée (eau + sels minéraux + substances organiques).
θ = %eau = [(MF – MS)/MF]*100 ; MS = Matière Sèche ; MF = Matière Fraîche
Déficit en eau : D= (θm – θ)/θm ; θm = teneur maximum ; θ= teneur réelle.
La teneur en eau diminue avec l’âge de la plante, dans les graines, on trouve entre 5 et 10% d’eau.
BLes états et rôles de l’eau dans la plante.
On trouve l’eau sous deux états :
- L’eau libre : elle peut être en solution (dans les vacuoles ou les sèves), sous forme de vapeur (dans les méats, dans la chambre sous-stomatique).
- L’eau liée : elle peut être liée par la force osmotique, par la force capillaire (tensions superficielles), par les forces d’imbibition (force électrostatique ou colloïde).
Les colloïdes sont des macromolécules très hydrophiles, comme les argiles ou l’humus. L’eau de constitution, est l’eau intra-moléculaire qui fait intervenir des forces très énergétiques.
Le CAH est le complexe argilo-humique.

Le rôle du CAH :
- mécanique : il maintient la turgescence des cellules, donc le port des végétaux. Une perte d’eau entraîne un flétrissement.
- physiologique : dans le milieu réactionnel de la cellule, l’eau est le solvant des molécules organiques. Cette eau sert de véhicule aux aliments et permet donc la circulation des constituants de la sève.
C la diffusion et la pression osmotique.
Le passage de molécules dissoutes à travers la membrane (avec perméabilité spécifique) entraîne un phénomène passif. De part et d’autre de la membrane, on assiste à un équilibrage grâce à l’agitation thermique : le mélange devient homogène. Avec une membrane hémiperméable, les courants d’eau vont égaliser les états énergiques des deux côtés.
La pression osmotique d’une solution, est l’attraction exercée par cette solution sur les molécules d’eau, lorsqu’elle est séparée par une membrane hémiperméable. La pression est donnée par la loi de Vant’Hoff : p.v = nRT = (m/n)RT ↔ p=(R.N)/(V.T)=RTC
Osmose = endosmose (ext. → int.) + exosmose (int. → ext.).
D Variation de la teneur en eau dans la cellule.
L’eau est en grande partie dans la vacuole. L’eau exerce, grâce à la vacuole, au cytoplasme, au plasmodesme, une pression de turgescence sur la paroi. Les ions et molécules, dans la vacuole, lui confèrent une pression osmotique qui attire l’eau.
La cellule dans un milieu hypotonique : l’eau rentre dans la cellule qui devient turgescente à cause d’une variation de volume cellulaire.
La cellule dans un milieu hypertonique : l’eau sort et on assiste à une plasmolyse.
Potentiel de l’eau ↔ pression de succion (ψ)
Potentiel hydrostatique ↔ pression de turgescence (ψt)
Potentiel osmotique ↔ pression osmotique (ψv)

Il y a évolution ψ = ψv + ψt à l’entrée de l’eau dans la cellule. Le xylème est lignifié pour éviter l’écrasement. On constate des variations de diamètre sur une plante pendant la journée.
E Absorption de l’eau.
En un jour, la plante absorbe un volume d’eau équivalent à sa masse. Les plantes aquatiques absorbent l’eau sur toute leur surface. Les plantes supérieures ont leur absorption assurée par leurs poils absorbant (fixés sur les racines), et pour beaucoup d’entre elles, par des mycorhizes. C’est l’eau la moins liée au sol qui va être utilisée par la plante (on parle de liaisons osmotiques et capillaires).
F Le transport de l’eau.
L’eau et les sels minéraux sont transportés par les poils absorbants jusqu’à l’endoderme. Le cheminement se fait par la loi de l’osmose (phénomènes physiques). Il y a déplacement par voie apoplastique (à travers la paroi) et symplastique (à travers les vacuoles et le cytoplasme).
Au niveau de l’endoderme, la couche cellulaire subérifiée tangentiellement (bandes de Caspary) oblige un passage par voie symplastique.
Jusqu’au cylindre central la pression (poussé) est racinaire. C’est un processus nécessitant de l’énergie métabolique.
Au niveau du cylindre central, on a un cheminement vertical qui se fait dans le xylème. Au début, la pression racinaire, devient au fur et à mesure de l’élévation par un processus physique fondé sur l’existence d’un gradient de potentiel hydrique (différence de potentiel entre le sol et l’atmosphère). L’assimilation (ou aspiration) par les feuilles permet l’évaporation (phénomène transpiratoire). L’atmosphère a un potentiel hydrique très négatif et soutire en permanence l’eau de la plante, créant ainsi un flux transpiratoire. Cette aspiration entraîne une dépression dans les vaisseaux du xylème. C’est ce phénomène qui a entraîné sa lignification.
G Emission d’eau par la plante.
Pour qu’il y ait transpiration, il faut que ψ soit très négatif. La différence de potentiels dépend de l’humidité relative de l’air. Le problème pour les plantes des pays équatoriaux est que l’humidité relative est de 100%. Il n’y a donc pas de différence de potentiels, donc pas de transpiration. Le transport est réalisé par les stomates aquifères hydatodes qui expulsent de l’eau. Pour cette vaporisation, il y a un besoin d’énergie solaire (95 pour cent par les stomates, 5% par la cuticule).
H Les stomates.
Ce sont des cellules particulières situées dans les feuilles. Elles ont une forme réniforme et sont accompagnées de 2 grosses cellules (cellules compagnes ou annexes) : c’est l’ostiole (là où circulent les gaz). Souvent, on observe la présence d’une chambre sous-stomatique.
Remarque : chez les monocotylédones, les stomates se trouvent sur les 2 faces des feuilles.
La structure :
- La paroi a des épaisseurs différentes : la partie interne est épaisse et cutinisée (d’où sa rigidité) ; la partie externe est fine et cellulosique (flexibilité).
- Si la cellule est turgescente, l’ostiole s’ouvre sinon elle reste fermée. C’est le mouvement de l’eau qui commande l’ouverture (K+, Cl–, malate2-).
- La mesure du potentiel membranaire met en évidence sur le plasmodesme, des pompes à protons de type ATPase qui interviennent en contrôlant la polarisation membranaire par l’expulsion de protons (besoin d’énergie).
On a mis en évidence des canaux ioniques sur la membrane, qui permettent aux ions de passer. Il l’énergie de la photosynthèse sert à faire sortir l’eau.
Remarque : les stomates à son ont les seules cellules épidermiques à posséder de la chlorophylle.
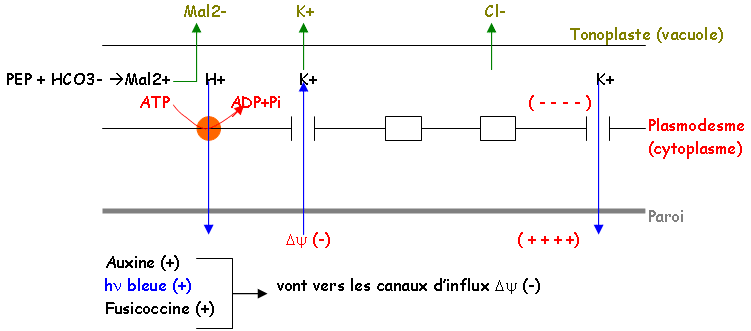
L’acide abscissique (-) (hormone) va vers les canaux d’efflux Δψ(+).
Souvent, les stomates son ont ouvert le jour (sauf chez les CAM). De plus, une faible concentration en CO2 dans la chambre sous-stomatique entraîne l’ouverture de l’ostiole.
H Circulation des sèves.
Cette différence entre la partie aérienne et le sol fait monter la sève brute. La sève élaborée est chargée de molécules organiques (dans le liber). Cette sève est transportée bidirectionnellement, du bas vers le haut et inversement, pour atteindre toutes les parties de la plante. Cette circulation se fait avec une pression positive : on parle de flux de pression. Cette sève contient de 50 à 300 g de matière sèche par litre.
La vitesse de flux.
La vitesse de la sève élaborée est inférieure au flux transpiratoire (de la sève brute). Malgré cela, le flux de sève élaborée ne se fait pas par simple diffusion. Actuellement, ce mécanisme de circulation de la sève élaborée est mal connu. La théorie actuelle est celle ne demande de l’énergie : flux volumique sous pression (ou flux de masse) : c’est la théorie de Münch. Le chargement des tubes criblés demande de l’énergie : ce sont des potentiels négatifs.
ψ = ψp + ψπ + ψh = P-(Ρ.g.z). ψh : C’est le potentiel gravitationnel, il dépend de la hauteur où l’on se trouve. Au niveau du sol, on a ψh=0.
Le phloëme : la sève élaborée s’est déchargée des solutés (ψ diminue). Il y a attirance du liber vers le xylème. La sève brute se charge en eau. Au sommet, La différence de pression entraîne le passage du haut du xylème vers le liber.
j’ai beaucoup aimé ce cour qui ma intruit